Энергообеспечение мышечной деятельности
Содержание:
- Этапы окислительного гликолиза. Фаза 1.
- Распространение и значение
- Суммарный выход гликолиза
- Гликолиз
- Бескислородное окисление глюкозы включает два этапа
- Примечания и ссылки
- Что такое гликолиз
- Фаза 2. Синтез Адезинтрифосфата
- 6-я ступень – окисление глицеральдегид-3-фосфата
- 7-я ступень. Перемещение фосфатной группы с 1,3-дифосфоглицерата на адезиндифосфат
- 8-я ступень. Перенесение фосфорильной группы с 3-фосфоглицерата.
- 9-я ступень. Выделение воды из 2-фосфоглицерата
- 10-я и последняя ступень. Перенос фосфатного остатка с ФЕП на АДФ
Этапы окислительного гликолиза. Фаза 1.
В аэробном гликолизе выделяются 2 фазы.
Первая фаза – подготовительная. В ней глюкоза вступает в реакцию с 2 молекулами АТФ. Эта фаза состоит из 5 последовательных ступеней биохимических реакций.
1-я ступень. Фосфорилирование глюкозы
Фосфорилирование, то есть процесс переноса остатков фосфорной кислоты в первой и последующих реакциях производится за счет молекул адезинтрифосфорной кислоты.
В первой ступени остатки фосфорной кислоты из молекул адезинтрифосфата переносятся в молекулярную структуру глюкозы. В ходе процесса получается глюкозо-6-фосфат. В качестве катализатора в процессе выступает гексокиназа, ускоряющая процесс с помощью ионов магния, выступающих в качестве кофактора. Ионы магния задействованы и в других реакциях гликолиза.

2-я ступень. Образование изомера глюкозо-6-фосфата

На 2-й ступени происходит изомеризация глюкозо-6-фосфата во фруктозу-6-фосфат.
Изомеризация – образование веществ, имеющих одинаковый вес, состав химических элементов, но обладающих разными свойствами вследствие различного расположения атомов в молекуле. Изомеризация веществ осуществляется под действием внешних условий: давления, температур, катализаторов.
В данном случае процесс осуществляется под действием катализатора фосфоглюкозоизомеразы при участии ионов Mg+.
3-я ступень. Фосфорилирование фруктозо-6-фосфата
На данной ступени происходит присоединение фосфорильной группы за счет АТФ. Процесс осуществляется при участии фермента фосфофруктокиназа-1. Этот фермент и предназначен только для участия в гидролизе. В результате реакции получаются фруктозо-1,6-бисфосфат и нуклеотид адезинтрифосфат.

АТФ – адезинтрифосфат, уникальный источник энергии в живом организме. Представляет собой довольно сложную и громоздкую молекулу, состоящую из углеводородных, гидроксильных групп, азота и групп фосфорной кислоты с одной свободной связью, собранных в нескольких циклических и линейных структурах. Высвобождение энергии происходит в результате взаимодействия остатков фосфорной кислоты с водой. Гидролиз АТФ сопровождается образованием фосфорной кислоты и выделением 40-60 Дж энергии, которую организм затрачивает на свою жизнедеятельность.
Но прежде должно произойти фосфорилирование глюкозы за счет молекулы Адезинтрифосфата, то есть перенос остатка фосфорной кислоты в глюкозу.
4-я ступень. Распад фруктозо-1,6-дифосфата

В четвертой реакции фруктозо-1,6-дифосфат распадается на два новых вещества.
- Диоксиацетонфосфат,
- Глицеральд альдегид-3-фосфат.
В данном химическом процессе в качестве катализатора выступает альдолаза, фермент, участвующий в энергетическом обмене, и необходимый при диагностировании ряда заболеваний.
5-я ступень. Образование триозофосфатных изомеров
И, наконец, последний процесс – изомеризация триозофосфатов.

Глицеральд-3-фосфат продолжит участвовать в процессе аэробного гидролиза. А второй компонент – диоксиацетон фосфат при участии фермента триозофосфатизомеразы преобразуется в глицеральдегид-3-фосфат. Но трансформация эта – обратимая.
Распространение и значение
Гликолиз является почти универсальным для всех клеточных организмов путем катаболизма глюкозы. Вероятно, он возник на ранних этапах развития жизни на Земле, еще до того, как в атмосфере накопился кислород в результате деятельности фотосинтезирующих организмов. Последовательность гликолитических реакций — чрезвычайно эволюционно консервативный путь. Ферменты, которые принимают в нем участие, проявляют высокую степень гомологии как в аминокислотной последовательности, так и трехмерной структуре у позвоночных животных, дрожжей и растений. Как показало секвенирования ДНК геномов большого количества организмов, только в некоторых архей и паразитических бактерий отсутствуют один или несколько ферментов гликолиза, несмотря на то, что сам путь сохраняется, вероятно, в модифицированном виде.
В эукариотических клетках реакции гликолиза происходят в цитозоле. В большинстве таких клеток именно этот среди других метаболических путей занимает первое место по количеству атомов углерода, которые превращаются в нем. Для таких тканей млекопитающих, как мозг (за исключением условий голодания), мозговой слой почек, сперматозоиды, а также эритроциты, в которых полностью отсутствуют митохонондрии, гликолиз является единственным источником метаболической энергии. Для мышц в условиях очень большой нагрузки гликолиз выгоден не только потому, что дает возможность получить энергию при недостатке кислорода, а еще и потому, что он происходит очень быстро и обеспечивает синтез АТФ в 10,5 раз быстрее, чем аэробная окисления органических веществ . Также от гликолиза главным образом зависят растительные ткани, специализированные на хранении крахмала (например клубни картофеля), и водные растения, такие как настурция лекарственная.
Другими путями окисления глюкозы является пентозофосфатный путь и путь Энтнера-Дудорова. Последний является заменой гликолиза в некоторых грамотрицательных и, очень редко, грамположительных бактерий и имеет много общих черт с ним ферментов.
Суммарный выход гликолиза
| Изменение свободной энергии в реакциях гликолиза в эритроцитах | ||
|---|---|---|
| Реакция | ΔG 0 (кДж / моль) | ΔG (кДж / моль) |
| Глюкоза + АТФ → глюкозо-6-фосфат + АДФ | -16,7 | -33,4 |
| Глюкозо-6-фосфат фруктозо-6-фосфат | 1,7 | от 0 до 25 |
| Фруктозо-6-фосфат + АТФ → фруктозо-1,6-бисфосфат + АДФ | -14,2 | -22,2 |
| Фруктозо-1,6-бисфосфат глицеральдегид-3-фосфат + дигидроксиацетонфосфат | 28,3 | от -6 до 0 |
| Дигидроксиацетонфосфат глицеральдегид-3-фосфат | 7,5 | от 0 до 4 |
| Глицеральдегид-3-фосфат + Ф н + НАД + 1,3-бисфосфоглицерат + НАДH + H + | 6,3 | от -2 до 2 |
| 1,3-бисфосфоглицерат + АДФ 3-фосфоглицерат + АТФ | -18,8 | от 0 до 2 |
| 3-фосфоглицерат 2-фосфоглицерат | 4,4 | от 0 до 0,8 |
| 2-фосфоглицерат фосфоэнолпируват + H 2 O | 7,5 | от 0 до 3,3 |
| Фосфоэнолпируват + АДФ → пируват + АТФ | -31,4 | -16,7 |
Общее уравнение гликолиза имеет следующий вид:
Глюкоза + 2Ф н + 2АДФ + 2НАД + → 2 пируват + 2АТФ + 2НАДН + 2Н + + 2Н 2 О.
Суммарное количество энергии, выделяющейся в процессе расщепления глюкозы до пирувата составляет 146 кДж / моль, на синтез двух молекул АТФ расходуется 61 кДж / моль, остальные 85 кДж / моль энергии превращается в тепло.
При полном окислении глюкозы до углекислого газа и воды выделяется 2840 кДж / моль, если сравнить это значение с общим выходом екзергоничних реакций гликолиза (146 кДж / моль), то становится понятно, что 95% энергии глюкозы остается «заключенной» в молекулах пирувата. Хотя реакции гликолиза являются универсальными почти для всех организмов, дальнейшая судьба его продуктов — пирувата и НАДН — отличается в разных живых существ и зависит от условий.
В аэробных организмов при достаточной концентрации кислорода НАД + регенерируется путем передачи электронов в дыхательная цепь переноса электронов, который в эукариот расположен во внутренней мембране митохондрий. Конечным акцептором электронов при этом является кислород. Пируват подвергается окислительного декарбоксилирования, превращается в ацетил-КоА и поступает в цикл Кребса, где происходит его дальнейшее окисление. Отщеплений электроны также попадают в дыхательная цепь переноса электронов.
С другой стороны, в анаэробных условиях восстановлен НАДH не может передать свои электроны на кислород, поэтому он переносит их или непосредственно назад на молекулу пирувата, как в процессе молочнокислого брожения, или на определенные продукты его превращения, например в ацетальдегид в случае спритового брожения. Анаэробный метаболизм глюкозы дает значительно меньше энергии, чем аэробный.
Гликолиз
Анаэробное превращение глюкозы локализуется в цитозоле и включает два этапа из 11 ферментативных реакций.
Первый этап гликолиза
Первый этап гликолиза – подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.
Первая реакция гликолиза сводится к превращению глюкозы в реакционно-способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода. Эта реакция является первой в любом превращении глюкозы, катализируется гексокиназой.
Вторая реакция необходима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования (фермент глюкозофосфат-изомераза). В результате образуется фруктозо-6-фосфат.
Третья реакция – фермент фосфофруктокиназа фосфорилирует фруктозо-6-фосфат с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. Эта реакция является главной в регуляции скорости гликолиза.
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных триоз-изомеров – альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакция подготовительного этапа – переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга при участии триозофосфатизомеразы. Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, определяет дальнейшую судьбу глюкозы:
- при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза,
- при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний отправляется на синтез глицеррол-3-фосфата для образования фосфатидной кислоты и далее тривцилглицеролов.
Второй этап гликолиза
Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
Шестая реакция гликолиза (фермент глицеральдегидфосфат-дегидрогеназа) – окисление глицеральдегидфосфата до кислоты и присоединение к ней фосфорной кислоты приводит к образованию макроэргического соединения 1,3-дифосфоглицериновой кислоты и НАДН.
В седьмой реакции (фермент фосфоглицераткиназа) энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ. Реакция получила дополнительное название – реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (от субстрата реакции) в отличие от окислительного фосфорилирования (использование энергии электрохимического градиента ионов водорода на мембране митохондрий).
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы изомеризуется в 2-фосфоглицерат.
Девятая реакция – фермент енолаза отрывает молекулу воды от 2-фосфоглицериновой кислоты и приводит к образованию макроэргической фосфоэфирной связи в составе фосфоенолпирувата.
Десятая реакция гликолиза – еще одна реакция субстратного фосфорилирования – заключается в переносе пируваткиназой макроэргического фосфата с фосфоенолпирувата на АДФ с образованием пировиноградной кислоты и АТФ.
Последняя реакция бескислородного окисления глюкозы, одиннадцатая – образование молочной кислоты из пирувата под действием лактатдегидрогеназы
Важно то, что эта реакция осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие кислорода не может окисляться в митохондриях
| У плода и детей первых месяцев жизни преобладает анаэробный распад глюкозы, в связи с чем концентрация молочной кислоты в крови у них выше чем у взрослых. |
| При наличии кислорода пировиноградная кислота переходит в митохондрию и превращается в ацетил-S-КоА. |
Бескислородное окисление глюкозы включает два этапа
Анаэробное превращение глюкозы локализуется в цитозоле и включает два этапа из 11 ферментативных реакций.
Первый этап гликолиза
Первый этап гликолиза — подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.
Первая реакция гликолиза сводится к превращению глюкозы в реакционно-способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода. Эта реакция является первой в любом превращении глюкозы, катализируется гексокиназой.
Вторая реакция необходима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования (фермент изомераза). В результате образуется фруктозо-6-фосфат.
Третья реакция — фермент фосфофруктокиназа фосфорилирует фруктозо-6-фосфат с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. Эта реакция является главной в регуляции скорости гликолиза.
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных триоз-изомеров — альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакция подготовительного этапа — переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга при участии триозофосфатизомеразы. Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97 %, доля глицеральдегидфосфата — 3 %.
Эта реакция, при всей ее простоте, определяет дальнейшую судьбу глюкозы:
- при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза:
- при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний отправляется на синтез жиров.
Второй этап гликолиза
Второй этап гликолиза — это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
Шестая реакция гликолиза (фермент глицеральдегидфосфат-дегидрогеназа) — окисление глицеральдегидфосфата и присоединение к нему фосфорной кислоты приводит к образованию макроэргического соединения 1,3-дифосфоглицериновой кислоты и НАДН.
В седьмой реакции (фермент фосфоглицераткиназа) энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ. Реакция получила дополнительное название — реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (от субстрата реакции) в отличие от окислительного фосфорилирования (от электрохимического градиента ионов водорода на мембране митохондрий).
Восьмая реакция — синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы изомеризуется в 2-фосфоглицерат.
Девятая реакция — фермент енолаза отрывает молекулу воды от 2-фосфоглицериновой кислоты и приводит к образованию макроэргической фосфоэфирной связи в составе фосфоенолпирувата.
Десятая реакция гликолиза — еще одна реакция субстратного фосфорилирования — заключается в переносе пируваткиназой макроэргического фосфата с фосфоенолпирувата на АДФ и образовании пировиноградной кислоты.
Последняя реакция бескислородного окисления глюкозы, одиннадцатая — образование молочной кислоты из пирувата под действием лактатдегидрогеназы
Важно то, что эта реакция осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие кислорода не может окисляться в митохондриях
У плода и детей первых месяцев жизни преобладает анаэробный распад глюкозы, в связи с чем концентрация молочной кислоты в крови у них выше чем у взрослых.
Примечания и ссылки
- Стивен Д. Антон , Keelin Moehl , Уильям Т. Донахью и Krisztina Marosi , « Подавать метаболический переключатель: Понимание и применение Польза для здоровья Голодание », Ожирение (. Silver Spring, MD) , Vol. 26, п о 2февраль 2018, стр. 254–268
- ↑ и (in) Реджинальд Х. Гарретт и Чарльз М. Гришем , Биохимия , Wadsworth Publishing Co Inc.,2012 г., 5- е изд. , 1280 с.
- Паскаль Риберо-Гайон , Дени Дюбурдье , Бернар Донеш и Алин Лонво , Договор об энологии , т. 1: Микробиология вина. Виноделие , Данод,3 октября 2012 г., 6- е изд. ( ISBN 978-2-10-058234-1 и 2-10-058234-8 )
-
(in) HS Seeholzer, A. Jaworowski, IA Rose , Энолпируват: химическое определение как промежуточное соединение пируваткиназы , Biochemistry , vol. 30, п о 3,
22 января 1991 г., стр. 727-732 -
Джордж Б. Джонсон , Джонатан Б. Лосос , Питер Х. Рэйвен и Сьюзан С. Сингер ( перевод с англ.), Биология: роскошная версия , Брюссель / Париж, Superior De Boeck
15 ноября 2009 г., 1406 с. ( ISBN 978-2-8041-6638-0 ) -
Жозеф-Пьер Гиро , Пищевая микробиология , Dunod,
18 сентября 2012 г., 2- е изд. ( ISBN 978-2-10-057008-9 и 2-10-057008-0 )
Что такое гликолиз
В общих чертах, гликолиз — это процесс, который происходит в цитоплазме всех клеток: от низших бактерий до высших млекопитающих — людей. Для некоторых это единственный метаболический путь доставки энергии клетке — например, для бактерий, для других же это метаболический путь, в который «сливаются» и другие метаболиты — белки и липиды.
Гликолиз — это основной путь расщепления глюкозы в нашем организме, основное биологическое значениеэтого процесса заключается в получении энергии и других продуктов обмена.
 Гликолиз происходит как в аэробных, так и в анаэробных условиях. Процесс в анаэробных условиях (недостаток кислорода) называется анаэробным гликолизом — конечным продуктом лактата и ферментации в дрожжах — алкогольной или молочнокислой ферментации.
Гликолиз происходит как в аэробных, так и в анаэробных условиях. Процесс в анаэробных условиях (недостаток кислорода) называется анаэробным гликолизом — конечным продуктом лактата и ферментации в дрожжах — алкогольной или молочнокислой ферментации.
При наличии кислорода этот процесс называется аэробным гликолизом — конечным продуктом выступают CO2 и H2O,
Гликолиз представляет собой серию реакций, в которых одна молекула глюкозы распадается на две молекулы пирувата. Но так происходит не всегда, в случаях когда доступ кислорода в этой реакции затруднен, проще говоря, в анаэробных условиях, то пируват превращается в лактат или спирт.
Механизмы анаэробного метаболизма: гликолиз в анаэробных условиях
В анаэробных условиях O2 отсутствует. Отсутствие конечного субстрата O2 ингибирует активность дыхательных структур (до этого они неактивны), и таким образом конечным продуктом распада глюкозы в анаэробных условиях является лактат,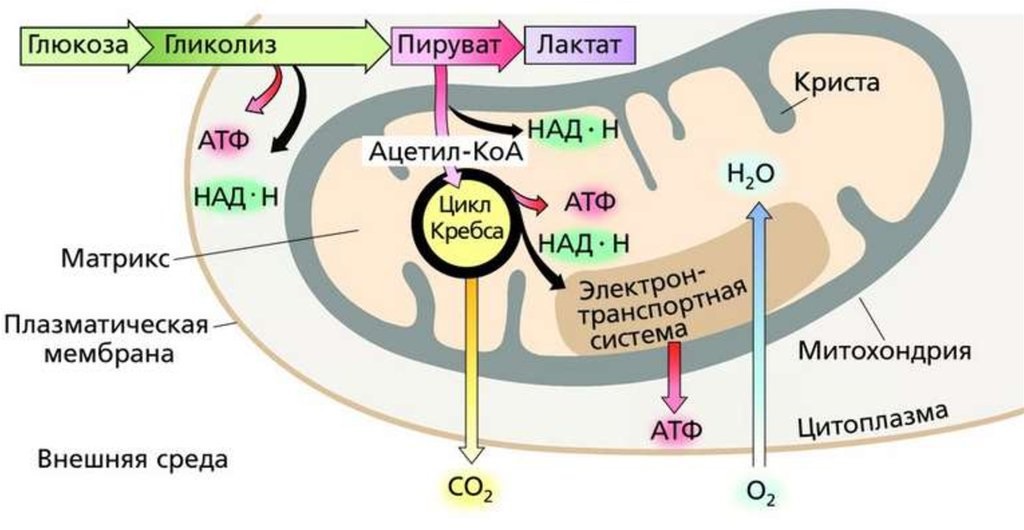
Анаэробный распад глюкозы в физиологических условиях наблюдается в эритроцитах (без митохондрий) и в сильно сокращающихся мышцах (недостаточное кровоснабжение кислородом), это наблюдается при сильной физической работе — присутствует истощение O2 в мышечных клетках (в форме оксимиоглобина).
Лактат в мышечных клетках приводит к мышечной лихорадке, которая преодолевается неинтенсивной мышечной активностью, направленной на полное расщепление лактата в мышцах, до CO2 , H2O и H2 в результате аэробного гликолиза.
При патологических состояниях и онкологических заболеваниях, у многих людей наблюдается так называемый эффект Пастера, при котором клетки из-за уменьшенного поступления крови или кислорода переключаются на анаэробный распад глюкозы, связанный с повышенным ее потреблением (повышенная экспрессия трансмиттеров глюкозы GLUT1 ) и гипогликемией.
Анаэробный гликолиз, как при физиологических, так и при патологических состояниях, характеризуется повышенным уровнем конечного продукта в результате расщепления его лактата в плазме крови — лактатемией, что может привести к ацидозу.
Дальнейшее расщепление лактата
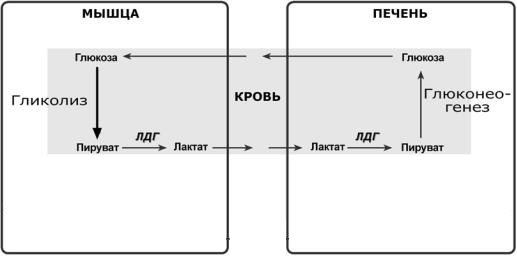
Лактат метаболизируется из печени в пируват, катализируя реакцию в обратном направлении, и глюкоза синтезируется пируватом с использованием ферментов глюконеогенеза в печени через цикл Кори (лактатный цикл).
Вот в этой схеме и кроется ответ на вопрос о том, как влияет анаэробный гликолиз на производство энергии, ведь мы видим, что наша молочная кислота, образовавшаяся в мышцах, через кровь попадает в печень и участвует в процессе глюконеогенеза — создания глюкозы из неуглеводных продуктов.
Фаза 2. Синтез Адезинтрифосфата
В данной фазе гликолиза будет аккумулироваться в виде АТФ биохимическая энергия. Адезинтрифосфат образуется из адезиндифосфата за счет фосфорилирования. А также образуется НАДН.
Аббревиатура НАДН имеет очень сложную и труднозапоминаемую для неспециалиста расшифровку – Никотинамидадениндинуклеотид. НАДН – это кофермент, небелковое соединение, участвующее в химических процессах живой клетки. Он существует в двух формах:
- окисленной (NAD+, NADox);
- восстановленной (NADH, NADred).
В обмене веществ NAD принимает участие в окислительно-восстановительных реакциях транспортируя электроны из одного химического процесса в другой. Отдавая, или принимая электрон, молекула преобразуется из NAD+ в NADH, и наоборот. В живом организме НАД вырабатывается из триптофана или аспартата аминокислот.
Две микрочастицы глицеральдегид-3-фосфата подвергаются реакциям, в ходе которых образуется пируват, и 4 молекулы АТФ. Но конечный выход адезинтрифосфата составит 2 молекулы, поскольку две затрачены в подготовительной фазе. Процесс продолжается.
6-я ступень – окисление глицеральдегид-3-фосфата
В данной реакции происходит окисление и фосфорилирование глицеральдегид-3-фосфата. В итоге получается 1,3-дифосфоглицериновая кислота. В ускорении реакции участвует глицеральдегид-3-фосфатдегидрогеназа

Реакция происходит при участии энергии, полученной извне, поэтому она называется эндергонической. Такие реакции протекают параллельно с экзергоническими, то есть выделяющими, отдающими энергию. В данном случае такой реакцией служит следующий процесс.
7-я ступень. Перемещение фосфатной группы с 1,3-дифосфоглицерата на адезиндифосфат

В этой промежуточной реакции фосфорильная группа переносится фосфоглицераткиназой с 1,3-дифосфоглицерата на адезиндифосфат. В итоге получаются 3-фосфоглицерат и АТФ.
Фермент фосфоглицераткиназа приобрел свое название за способность катализировать реакции в обоих направлениях. Этот фермент также транспортирует фосфатный остаток с адезинтрифосфата на 3-фосфоглицерат.
6-я и 7-я реакции часто рассматриваются как единый процесс . 1,3-дифосфоглицерат в нем рассматривается как промежуточный продукт. Вместе 6-я и 7-я реакции выглядят так:
Глицеральдегид-3-фосфат+ADP+Pi +NAD+⇌3 -фосфоглицерат+ATP+NADH+Н+,ΔG′о = −12,2 кДж/моль.
И суммарно эти 2 процесса освобождают часть энергии.
8-я ступень. Перенесение фосфорильной группы с 3-фосфоглицерата.

Получение 2-фосфоглицерата – процесс обратимый, происходит под каталитическим действием фермент фосфоглицератмутазы. Фосфорильная группа переносится с двухвалентного атома углерода 3-фосфоглицерата на трехвалентный атом 2-фосфоглицерата, в итоге образуется 2-фосфоглицериновая кислота. Реакция проходит при участи положительно заряженных ионов магния.
9-я ступень. Выделение воды из 2-фосфоглицерата
Эта реакция в своей сути является второй реакцией расщепления глюкозы (первой была реакция 6-й ступени). В ней фермент фосфопируватгидратаза стимулирует отщепление воды от атома С, то есть процесс элиминирования из молекулы 2-фосфоглицерата и образование фосфоенолпирувата (фосфоенолпировиноградной кислоты).

10-я и последняя ступень. Перенос фосфатного остатка с ФЕП на АДФ
В заключительной реакции гликолиза задействованы коферменты – калий, магний и марганец, в качестве катализатора выступает фермент пируваткиназа.

Преобразование енольной формы пировиноградной кислоты в кето-форму является обратимым процессом, и в клетках присутствуют оба изомера. Процесс перехода изометрических веществ из одного в другой называется таутомеризацией.